مگس سرکه
مگس سرکه یا مگس میوه (نام علمی: Drosophila melanogaster)، گونهای مگس از راستهٔ دوبالان است.[۱]
| مگس سرکه | |
|---|---|

| |
| مگس سرکهٔ نر | |
| ردهبندی علمی | |
| حوزه: | یوکاریا |
| فرمانرو: | جانوران |
| شاخه: | بندپایان |
| رده: | حشرات |
| راسته: | دوبالان |
| تیره: | مگسهای میوه |
| سرده: | دروزوفیلا |
| زیرسرده: | Sophophora |
| Species group: | melanogaster group |
| Species subgroup: | melanogaster subgroup |
| Species complex: | melanogaster complex |
| نام دوبخشی | |
| Drosophila melanogaster | |
آرایهشناسی
ویرایشمگس سرکه، گونهای از سردهٔ دروزوفیلا و از ردهٔ حشرات (راستهٔ آرایهشناسی دوبالان) در خانوادهٔ مگسهای میوه است. این گونه، با نام «مگس میوهٔ معمولی» نیز شناخته میشود.[۲]
بهکارگیری در آزمایشها
ویرایشدر آغاز با پیشنهاد کاربرد مگس سرکه بهعنوان یک جاندار مدل از چارلز دابلیو وودوُرث، این گونه بهطور گستردهای در پژوهشهای زیستشناسی، در مطالعات ژنتیک، فیزیولوژی، بیماریزایی میکروبی و فرگشت پیشینهٔ زندگی بهکار برده شد.[۳]
ریختشناسی
ویرایش
مگسهای میوهٔ نوع وحشی، به رنگ زرد-قهوهای با چشمان قرمز آجری و حلقههای سیاه عرضی در سراسر شکم هستند. آنها دودیسی جنسی را نشان میدهند: مادهها نزدیک به ۲٫۵ میلیمتر (۰٫۰۹۸ اینچ) طول دارند و نرها کمی کوچکتر با پشت تیرهتر هستند. نرها به آسانی براساس تفاوتهای رنگ، از مادهها متمایز میشوند؛ همچنین با یک تکهٔ متمایز سیاه (کمتر دیدهشده در مگسهای نوظهور) در سطح شکمی (نگارهٔ ۱ را ببینید) و شانههای جنسی (ردیفی از موهای تیره بر روی قوزک نخستین پا).[۴]
چرخهٔ زندگی
ویرایش
پس از لقاح و پدیدآمدن زیگوت، مراحل رشد و نمو رویانی در داخل غشاهای تخم انجام میشود و لارو از تخم خارج میشود. سپس با تغذیه و رشد خود، در نهایت به شفیره تبدیل میشود. لاروها سفید رنگ و بندبند، بدون پا و زوائد بدنی و در ناحیهٔ سر، دارای قلابهای آروارهای به رنگ سیاه هستند. در این مرحله، رشد حشره سریع است و غذای فراوانی میخورد ولی هنوز بال ندارد. پس از آن، شفیره تکامل پیدا کرده و حشرهٔ کامل یا بالغ ظاهر میشود.[۵]
تغذیه و رشد
ویرایشمگس سرکه، به بوی تخمیر و سرکه جلب میشود و مخمرهای عامل گندیدگی میوهها را با خود بر روی میوههای گندیده یا رسیده حمل میکند و در آنجا کشت میدهد تا بستری برای تخمریزی فراهم شود. مگسهای کامل و لاروهای آنها، در محیطهای اسیدی و تخمیری زندگی میکنند و در محل تغذیهشان معمولاً میکروارگانیسمهایی مثل باکتریها و قارچها نفوذ میکند و تکثیر مییابد.
مگس کامل از ریسههای روی کمپوست تغذیه میکند، در داخل کمپوست تخمریزی مینماید و پس از طی مراحل بالا، لارو را بهوجود میآورد. لاروها شروع به تغذیه از ریسههای داخل کمپوست میکنند و اگر بر روی کمپوست قارچی وجود داشته باشد، از طریق پایه وارد کلاهک میشوند و کانالها و دالانهای فراوانی را در آن ایجاد خواهند کرد.[۵]
بستگی به دمای پیرامون
ویرایشطول مراحل چرخهٔ زندگی مگس سرکه، تحت تأثیر دمای محیط، متغیر است؛ بهطوریکه در دمای ۲۰ درجهٔ سانتیگراد، متوسط دورهٔ تخم و لارو، ۸ روز است و در دمای ۲۵ درجه، این مدت به ۵ روز کاهش مییابد. در دمای ۳۰ درجهٔ سانتیگراد در اثر تنش دمایی، متوسط مدت زمان تخم تا جاندار بالغ، ۱۱ روز است؛ در دمای ۲۸ درجه، مدت زمان تکوین به ۷ روز کاهش مییابد (بهترین دما)؛ این در حالی است که چرخهٔ تکوین در دماهای ۲۵، ۱۸ و ۱۲ درجه -در تنش سرما- (به ترتیب) تا ۸٫۵ روز، ۱۹ روز و بیش از ۵۰ روز افزایش خواهد یافت.
هرگاه دمای محیط از دمای بهینه (۲۵ تا ۲۸ درجهٔ سانتیگراد) پایین بیاید، قدرت تولیدمثل کم میشود؛ بهگونهای که سرانجام، در زیر ۱۶ درجهٔ سانتیگراد، باعث از بین رفتن مگسها میگردد. همچنین اگر دمای محیط بالاتر از ۳۰ درجهٔ سانتیگراد باشد، در پایان منجر به عقیم شدن حشرهٔ نر و از بین رفتن آنها خواهد شد.[۶]
ویژگیهای مدل
ویرایش
توماس هانت مورگان (زیستشناس برندهٔ جایزهٔ نوبل فیزیولوژی یا پزشکی سال ۱۹۳۳ م)، مگس سرکه را بهدلیل داشتن ویژگیهای زیادی برای بررسیهای ژنتیکی مورد استفاده قرار داد که برخی از آنها عبارتاند از:
- تعداد کروموزومها: این حشره، 2n کروموزومی است و تعداد کروموزومهایش کم (2n=۸) است. بررسی یک ژن یا یک جهش در ۸ کروموزوم، آسانتر است (نگارهٔ ۳ را ببینید).
- جنسیت مشخّص: تعیین جنسیت مگس سرکه آسان است. ژنوتیپ ماده xx و ژنوتیپ نر xy است؛ پس تعیین جنسیت با هتروگامت، یعنی نر است (نگارهٔ ۳ را ببینید). با دیدن علائم ظاهری نیز به سادگی میتوان تعیین جنسیت کرد.
- چرخهٔ زندگی کوتاه: یک دورهٔ زندگی مگس سرکه، در حدود ۱۰ روز طول میکشد.
- نرخ بالای زادآوری: تعداد فرزندان در هر باروری، ۴۰۰ تا ۵۰۰ زاده (از ۸۰۰ تا ۲۰۰۰ تخم) است.[۲]
- نبود مشکلات اخلاقی: کار با یک حشره در آزمایشهای ژنتیکی، با محدودیت اخلاقی مواجه نیست.
- دیانای دو رشتهای: داشتن دیانای دورشتهای در جاندار مورد آزمایش (همچون مگس سرکه)، از موضوعات دارای اهمیت است.
- کروموزوم پلیتن (ساختار کروموزومی غولپیکر): پلیتن در دورهٔ لاروی قابل استخراج است. امتیاز کروموزوم پلیتن این است که بررسی یک حذف یا یک جهش ژنی در آن، بسیار راحتتر است.[۷][۸]
نمو (سلولی و ملکولی)
ویرایش
تعیینکنندهها ترکیبهای ویژهای هستند که در برخی مجموعههای یاختهای در مراحل جنینی یافت میشوند و عاملی برای تمایز اندامها و بخشهای خاصی از بدن هستند. در بدن لارو مگس سرکه نواحی به نام صفحههای زایا وجود دارند. هر جفت صفحهٔ روبهروی هم، تعیینکنندهٔ اندام ویژهای مثل بالها، پاها و هالترها هستند (نگارهٔ ۴ را ببینید).
برهمکنشهای یاختهها بر تمایز یاختهای اثر زیادی دارد. به تدریج که نمو پیشرفت میکند، جنبشها و برهمکنشهای یاختهها بیشتر میشود. برای مثال، در مرحلهٔ گاسترولا، جابهجایی یاختهها موجب درونبرگشتگی رویان میشود و به دنبال آن، تعداد زیادی از یاختهها به درون برگشتگی (نوتوکورد)، اکتودرم را برای تبدیل به عصب، القا میکنند.
در جانداران پریاختهای یاختههای بافتهای متفاوت میتوانند به حالت موزون همکاری کنند. یاختهها میتوانند برهم اثر بگذارند و این برهمکنشها میتواند در فواصل نزدیک یا دور انجام شود. برهمکنشهای در فواصل دور مثل القاهای جنینی با نشر مواد شیمیایی برقرار میشوند. در بیشتر موارد، برهمکنشها در فواصل نزدیک یا در نتیجهٔ تماس یاختهها انجام میشوند. اهمیت ویژگیهای بازشناسی سطح یاختهها در برخی موارد و از جمله هنگام نمو دستگاه عصبی بهخوبی مشخص میشود. در این هنگام میلیونها نورون بایستی همتای مناسب خود را بیابند و با آن اتصالهای سیناپسی را که برای جریانهای عصبی پیچیده لازم است برقرار کنند.
با وجود پژوهشهای زیاد، ساختار شیمیایی مواد قابل انتشار که در القای جنینی دخالت دارند، هنوز به خوبی شناخته نشده است. القاکنندهها و تعیینکنندهها، نمونههایی از این ترکیبات هستند.
القاکنندهها ترکیبات پیچیدهای هستند که اغلب در مراحل ابتدایی نمو جنینی از لایههای سهگانهٔ اکتودرمی، مزودرمی و آندودرمی ترشح میشوند و موجب القای تشکیل اندام خاصی میگردند. برای مثال تشکیل لولهٔ عصبی از اکتودرم به وجود لایهٔ مزودرمی زیر آن نیاز دارد (تجربهٔ اسپمن) زیرا القاکنندهها از مزودرم به اکتودرم میرسند. پیوند این صفحهها به بدن جانور بالغ، موجب بروز اندامی که تعیینکنندهٔ آن نمیشود، اما اگر آنها را به بدن لارو مگس سرکه، در محلی غیر از جایگاه اصلی پیوند بزنیم، اندام خاص خود را تولید خواهند کرد. این توانایی تا سالهای طولانی در صفحههای زایا حفظ میشود (لایههای زایندهٔ جنینی -اکتودرم، مزودرم و آندودرم- حد واسط حالت پرتوان و یکتوان هستند؛ پس میگوییم چندتوان هستند). تجربههای انجام شده نشان دادهاند که اگر این صفحههای زایا را تا نسلهای متوالی (۱۸۰۰ نسل، طی ۹ سال) به مگس سرکهٔ بالغ پیوند بزنیم و سپس آنها را به لارو مگس سرکه پیوند کنیم، اندام ویژهٔ وابسته به خود را به وجود میآورند.[۹]
ژنتیک
ویرایش
کروموزوم پلیتن
ویرایشکروموزومهای پلیتن، کروموزومهای بزرگتر از اندازهٔ ویژهای هستند که از کروموزومهای استاندارد پرورش مییابند و عموماً در غدد بزاقی مگس سرکه یافت میشوند. سلولهای تخصصیافتهای، تحت تأثیر رونویسیهای پیدرپیِ دیانای بدون تقسیم سلول (اندومیتوز)، یک کروموزوم غولآسای پلیتن را میسازند تا حجم سلول را افزایش دهند. ساختار کروموزومهای پلیتن، در هنگام چندین دور رونویسی پیدرپیِ دیانای، کروماتیدهای خواهری زیادی را میسازد.[۱۰][۱۱]
ویژگیهای ژنی و کروموزومی
ویرایش

در ساختار کروموزوم پلیتن مگس سرکه، کروموزوم شمارهٔ ۴ و کروموزوم ایکس، بازو ندارد. کروموزومهای بزرگ شماره ۲ و ۳، متاسانتریک هستند (سانترومر در مرکز آنها قرار دارد)؛ در نتیجه، هر یک دارای دو بازوی راست و چپ هستند. کروموزومهای ۴ و ایکس، بازویی ندارند؛ چرا که تلوسانتریک هستند (سانترومر آنها در انتهای کروموزوم قرار دارد). ساختار کروموزوم پلیتن، هم در مگس سرکهٔ نر و هم در ماده وجود دارد، ولی در جنس ماده، کروموزوم Y وجود ندارد (نگارهٔ ۳ را ببینید). چرا که کروموزوم Y به صورت هتروکروماتینی است و در جنس نر دیده نمیشود. در بازوی R2، در قسمتی کروموزومهای همولوگ از هم جدا هستند. بررسیهای گوناگونی، این فاصله را در کروموزومهای دیگر (مانند L3) نیز نشان دادهاند (نگارهٔ ۵ را ببینید).
در مگس سرکه حدود ۲۰ ژن دارای هومیوباکس وجود دارد که حدود ۱۵تای آنها بررسی شدهاند و مشخص شده است که تمامی آنها قطعهبندی مگس سرکه را کنترل میکنند. تمام ژنهای هومیوباکس مگس سرکه جز یکی از آنها که در یاختههای خلفی قرار دارد و در ایجاد قطبیت پشتی-شکمی جنین مؤثر است، در دو مجموعهٔ مجزا به نام «Ant–C» و «Bx-C»، مجاور هم و بر روی بازوی راست کروموزوم ۳ قرار دارند (نگارهٔ ۶ را ببینید). برخی ژنهای دارای هومیوباکس نیز در قسمتهای دوردست ژنوم و در نواحی دیگری از آن (جز محل ۲ مجموعه) مشاهده شده است.[۱۲]
همانندی با ژنوم انسان
ویرایشحدود ۷۵ درصد از ژنهای شناخته شدهٔ بیماریهای انسانی، یک همتای قابل تشخیص در ژنوم مگسهای میوه دارد و ۵۰ درصد از توالیهای پروتئینی مگس، همولوگهای پستانداری دارد.[۱۳] یک پایگاه دادهٔ برخط (online database) به نام Homophila برای جستوجوی همتاهای ژنی بیماریهای انسانی در مگسها و برعکس، در دسترس است.[۱۴] مگس سرکه، بهعنوان یک مدل ژنتیکی برای بیماریهای مختلف انسانی، شامل بیماری پارکینسون، هانتینگتون، عدم تعادل مخچهای (SCA) و بیماری الزایمر استفاده میشود. مگس سرکه، همچنین برای مطالعهٔ سازوکارهای پیری اساسی و استرس اکسیداتیو، ایمنی، دیابت و سرطان، بهعلاوهٔ سوءمصرفهای دارویی مورد استفاده قرار گرفته است.
سازوکار مولکولی تمایززایی
ویرایشدر بیشتر پروکاریوتها، دیانای در اثر متیلاسیون در موقعیت ۵ سیتوزین، تغییراتی پیدا میکند. بیشتر ژنهای فعال یا دارای توان فعالیت، به این ترتیب متیله میشوند. اساس مولکولی این پدیده هنوز ناشناخته است. تصور میشود که گروههای متیل موجود در شیار بزرگ دیانای میتوانند بیان ژنها را با افزایش و کاهش میل ترکیبی دیانای با پروتئینهای تنظیمی مهارکننده یا فعالکننده، تنظیم کنند.
در طول فاز S، هنگامی که چنگال همانندسازی دیانای از میان دینوکلئوتید ۳َـCPGـ۵َ میگذرد، هر یک از مارپیچهای دختر بهطور موقت نیمهمتیله هستند (تنها رشتهٔ قدیمی متیله است). اگر هیچ راهی برای ترمیم این گروه متیله وجود نداشته باشد، به سرعت و طی زایشهای بعدی، گروه متیله در جمعیت کاهش مییابد، اما متیلاسیون به ارث میرسد. به نظر میرسد که یاختهٔ دارای یک آنزیم متیلاز دائمی، در یاختههای یوکاریوتی وجود دارد و نیز متیلاسیون توانایی تنظیم فعالیت ژنی را دارد، میتوان نتیجهگیری کرد که متیلاسیون در دینوکلئوتید ۳َـCPGـ۵َ، راهی برای پایداری حالت تمایز در یوکاریوتها است. اگر دیانای ژنومی در یاختههای جنسی یا در برخی از یاختههای بسیار ابتدایی جنینی بهطور خودبهخود به نوعی متیلاسیون برای جلوگیری از تمایز دست مییابد، پس متیلاسیون و دِمتیلاسیون در جایگاههای خاصی از کروموزوم میتواند در بیان برنامهریزی شدهٔ ژنها در طول تکوین نقشی ایفا نماید. مسئلهٔ اصلی در این مورد این است که متیلاسیون با دِمتیلاسیون، خود چگونه تنظیم میگردد. همهٔ متیلازهای دائمی یوکاریوتی که تاکنون شناخته شدهاند، در فعالیت خود، غیراختصاصی عمل میکنند. هر دینوکلئوتید CPG که در اختیار این آنزیمها قرار گیرد، متیله میشود. پذیرفته شده که عوامل دیگری مانند پروتئینهای تنظیمکننده که اتصالهای اختصاصی دارند یا تغییر ساختاری کروماتین، برای در دسترس قراردادن سیتوزینها عمل میکنند.[۷][۹]
تعیین جنسیت
ویرایشبا مشاهدهٔ ویژگیهای ظاهری مگس زیر میکروسکوپ نوری، میتوان به جنسیت آن پی برد.
* جنس نر: کوچکتر، تیرهتر (این ویژگی، خیلی تشخیصی نیست؛ یعنی همیشه نمیتوان بر پایهٔ رنگ بدن به آسانی نر و ماده را متمایز کرد)، دارای نوار تیرهٔ پهن در بند انتهایی بدن، دارای انتهای بدن بیضیشکل، دارای شکم ۵بندی و دارای نقطهای سیاه در پدیپالپ، (که شانهٔ جنسی نام دارد) است.
* جنس ماده: بزرگتر، روشنتر، دارای نوار تیرهٔ انتهایی نازکتر، دارای انتهای بدن نوکتیز و نیزهمانند، دارای شکم ۷بندی و فاقد شانهٔ جنسی است.[۴]
جهشیافتهها
ویرایشجهشهای ژنی در مگس سرکهٔ وحشی (wild type)، موجب زاده شدن مگس سرکهٔ جهشیافته (mutant) میشود. انواع گوناگونی از اثرات جهش ژنتیکی در این حشره شناسایی شده است و برای آزمایشهای ژنتیکی، بهطور تکی (مونوهیبریدیسم) یا دوتایی و بهصورت همزمان (دیهیبریدیسم) مورد استفاده قرار میگیرد؛ از جمله:
- دگرگونی بالها:
- [بسیار] بالکوتاه (Vestigial): برای اینکه صفت بالکوتاهی در مگس سرکه دیده شود، باید هر دو آلل ژن کدکنندهٔ صفت طول بال، جهشیافته (mutant) باشد؛ یعنی ژنوتیپ aa داشته باشد. مگس vestigial (بالکوتاه)، معمولاً نمیتواند پرواز کند و انتخاب طبیعی، آن را از محیط حذف میکند (بخش «نگارخانه» را ببینید). از آنجا که این جهشیافتهها به سن تولیدمثل نمیرسند و نمیتوانند ژن خود را به نسل بعد منتقل کنند، انتظار میرود که ژنوتیپ والدین، هتروزیگوت یا حامل (Aa or carrier) باشد.
- بالکوتاه مینیاتوری (Dumpy): جهش در ژن طول بال، میتواند منجر به پدید آمدن تیپ dumpy شود. همواره باید به تفاوتهای dumpy و vestigial توجه کرد. dumpy کمی بلندتر از vestigial و البته کوتاهتر از wild type است؛ همچنین حاشیهٔ بال در dumpy، صاف و بدون دندانه است، که در نوع vestigial، کنگرهدار و نامنظم است.
- دگرگونی رنگ بدن:
۱) بدن سیاه براق (Ebony): رنگ بدن مگس ebony در مقایسه با تیپ وحشی، بسیار تیرهتر و چشمها به رنگ قهوهای است و جوانترها دارای بدنی روشنتر از مسنها هستند. باید توجه داشت که در برخی موارد، فقط هالهای تیره در سطح پشتی بدن و از ناحیهٔ اتصال سر به سینه تا انتهای بدن وجود دارد که این مگسها نیز ebony محسوب میشوند. ژن ebony روی کروموزوم شمارهٔ ۳ مگس سرکه قرار دارد. جهش Recessive در ژن ebony، موجب تیرهشدن رنگ بدن و بالها میشود. رنگ بدن در حالت معمول (wild type)، زرد مایل به قهوهای است؛ ولی رنگ بدن نوع جهشیافتهٔ ebony، قهوهای تیره تا سیاه میشود (بخش «نگارخانه» را ببینید).
۲) بدن زرد روشن (Yellow): در این نوع جهشیافته، بدن به رنگ زرد روشن است که البته رنگ بدن، آیتم مناسبی برای تمایز yellow از wild type نیست. وجه تمایز yellow این است که در این نوع جهشیافته، بندهای بدن به صورت فراگمنت (تکهتکه) و مشخص وجود ندارد؛ خطوط تیره به فرم خیلی کمرنگ دیده میشود و به خوبی قابل تمایز از نوع وحشی است (بخش «نگارخانه» را ببینید).
- دگرگونی رنگ چشم (Eye color changes): رنگ چشم مگس سرکهٔ وحشی، قرمز است؛ اما در اثر جهش در ژن تعیینکنندهٔ رنگ چشم مگس، ممکن است چشمها به رنگ سفید، خاکستری، اسکارلت (قرمز شفاف)، قهوهای و… درآید (بخش «نگارخانه» را ببینید).
۱) چشم قرمزقهوهای (sepia): در sepia، مانند wild type، رنگدانههای قرمز و قهوهای، همزمان برای چشم بیان میشود؛ با این تفاوت که رنگدانهها منظم نیست (ناشی از بیان همتوان ژن) و حاشیهها در sepia قرمز و در مرکز بین قرمز و قهوهای دیده میشود (بخش «نگارخانه» را ببینید)؛ برخلاف wild type که اینچنین نیست (و ناشی از بیان حدواسط ژن است).
۲) چشم سفید (White): در این نوع جهشیافتهها، ژن هیچیک از دو رنگ بیان نمیشود؛ در واقع، هیچ رنگدانهای در چشم این جهشیافتهها نیست. گاهی ممکن است در مگسهایی با بدن تیره (مثل ebony)، چشمها به رنگ بدن دیده شود یا بهدلیل تیره بودن بدن، درست مشاهده نشود.[۱۵]
گامتزایی و نمو یاختههای جنسی
ویرایش
در مگس سرکه، سلولهای جنسی بدوی، به صورت دستهای از سلولهایی به نام سلولهای قطبی (Pole cells) در ناحیهٔ قطبی بلاستودرم به وجود میآیند. برخی از هستهها در نهمین تقسیم هستهای، به طرف ناحیهٔ عقبی مهاجرت میکنند و توسط پلاسم قطبی (Pole plasm) احاطه میشوند. پلاسم قطبی، مجموعهای از میتوکندریها، فیبریلها و دانههای قطبی (Pole granules) است. اگر هستههای سلول قطبی، توسط پلاسم قطبی احاطه نشود، سلولهای زاینده به وجود نمیآیند. یکی از ترکیبات پلاسم قطبی mRNAی، ژن (gcl (Germ cell-less است. ژن gcl، در سلولهای پرستار تخمدان بیان شده و mRNAی آنها به تخمک منتقل میشود. این mRNA در داخل تخمک، عقبیترین ناحیهٔ آن منتقل میگردد؛ این ناحیه، به پلاسم قطبی تبدیل میشود. این mRNA در مراحل اولیهٔ تسهیم، به پروتئین بیان میشود. تصور میشود که پروتئین حاصله از ژن gcl، وارد هسته شده و برای ایجاد سلول قطبی نیز ضروری است. در حشراتی که ژن gcl آنها جهش یافته باشد، سلولهای زاینده وجود ندارد.
علاوه بر پروتئین gcl، پروتئینهایی نظیر اسکار (Oskar)، نانوس (Nanos) و واسا (Vasa) نیز در پلاسم زایندهٔ برخی از گونهها وجود دارد که در ایجاد سلولهای زایندهٔ بدوی دخالت دارند.
گروهی دیگر از ترکیبات که در پلاسم زاینده وجود دارد، آرانای ریبوزومی-میتوکندریایی (mtr=ribosomal mitochondrial RNA) است که در ایجاد سلولهای جنسی بدوی دخالت دارد. در تخمکهای طبیعی حشرات، rRNAهای میتوکندریایی کوچک و بزرگ در بیرون میتوکندری و فقط در پلاسم قطبی جنینها در مرحلهٔ تسهیم وجود دارد که بهصورت دانههای قطبی دیده میشود. ترکیب دیگری که در پلاسم قطبی مگس سرکه و همچنین در دانههای قطبی وجود دارد، یک آرانای غیرقابل ترجمه به نام ترکیب دانهٔ قطبی (Polar granule component) است. عمل این ترکیب معلوم نیست؛ ولی در حشرات ترانسژنی که فاقد این ترکیب هستند، سلولهای زایندهٔ بدوی، قادر به مهاجرت به غدد جنسی نیستند.
طی جنینزایی مگس سرکه، سلولهای زایندهٔ بدوی، از قطب عقبی به غدد جنسی مهاجرت میکنند. اولین مرحله از این مهاجرت، به صورت غیرفعال است و طی آن ۳۰ تا ۴۰ سلول توسط حرکات گاسترولاسیون به لولهٔ گوارش عقبی جابهجا میشوند. در دومین مرحله، اندودرم لولهٔ گوارشی باعث شروع حرکت آمیبی در سلولهای جنسی بدوی میشود که از طریق انتهای کور بخش عقبی لولهٔ گوارش میانی، به مزودرم جداری مهاجرت میکنند. در سومین مرحله، سلولهای زایندهٔ بدوی به دو گروه تقسیم میشوند و هر یک از آنها در رابطه با جوانهٔ غدد جنسی در حال رشد قرار میگیرند. در چهارمین مرحله، سلولهای زایندهٔ بدوی به درون غدد جنسی مهاجرت میکنند. غدد جنسی از مزودرم جانبی به وجود میآیند. به نظر میرسد که محصول ژن وونن (Wunen) مسئول هدایت مهاجرت سلولهای زایندهٔ بدوی از اندودرم به مزودرم است. این پروتئین، درست قبل از مهاجرت PGC در اندودرم بیان میشود. پروتئین دیگری که در مهاجرت صحیح دخالت دارد، محصول ژن کلمبوس (Columbus) است. در آخرین مرحله، غدد جنسی در اطراف سلولهای زایندهٔ بدوی قرار میگیرند و به سلولهای زاینده اجازهٔ تقسیم میدهند و گامتها بالغ میشوند.[۱۶]
بهعنوان آفت
ویرایشمگس سرکه، معمولاً به دلیل گرایشش به تهاجم به منازل و ابنیهای که در آنها میوه یافت میشود، یک آفت کشاورزی در نظر گرفته میشود. مگسها ممکن است در خانهها، رستورانها، فروشگاهها و اماکن دیگر تجمع یابند.[۱۷] از بین بردن یک تهاجم، میتواند دشوار باشد؛ حتی زمانی که جمعیت بزرگسال (بالغ) هم نابود شده باشد، بهعنوان یک لارو، میتواند در میوهٔ مجاور، به بیرون آمدن از تخم ادامه دهد.
نگارخانه
ویرایش-
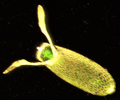 تخم مگس سرکه
تخم مگس سرکه -
 رنگهای گوناگون چشم و بدن در گونهٔ وحشی و گونههای جهشیافتهٔ مگس سرکه
رنگهای گوناگون چشم و بدن در گونهٔ وحشی و گونههای جهشیافتهٔ مگس سرکه -
 محیطهای کشت آزمایشگاهی مگس سرکه
محیطهای کشت آزمایشگاهی مگس سرکه -
 نمای جلو از مگس سرکه
نمای جلو از مگس سرکه -
 نمای کنار از یک مگس سرکهٔ جهشیافته
نمای کنار از یک مگس سرکهٔ جهشیافته -
 گونهای جهشیافته از مگس سرکه، روی یک گیاه
گونهای جهشیافته از مگس سرکه، روی یک گیاه -
 گونهای جهشیافته از مگس سرکه، روی یک گیاه
گونهای جهشیافته از مگس سرکه، روی یک گیاه



.jpg)
.jpg)


منابع
ویرایش- ↑ ۱٫۰ ۱٫۱ Meigen JW (1830). Systematische Beschreibung der bekannten europäischen zweiflügeligen Insekten. (Volume 6) (PDF) (به آلمانی). Schulz-Wundermann. Archived from the original (PDF) on 9 February 2012. Retrieved 3 December 2011.
- ↑ ۲٫۰ ۲٫۱ James H. Sang (2001-06-23). "Drosophila melanogaster: The Fruit Fly". In Eric C. R. Reeve (ed.). Encyclopedia of genetics. USA: Fitzroy Dearborn Publishers, I. p. 157. ISBN 978-1-884964-34-3. Retrieved 2009-07-01.
- ↑ Pierce, Benjamin A (2004). Genetics: A Conceptual Approach (2nd ed.). W. H. Freeman. ISBN 978-0-7167-8881-2.
- ↑ ۴٫۰ ۴٫۱ "FlyBase: A database of Drosophila genes and genomes". Genetics Society of America. 2009. Archived from the original on 15 August 2009. Retrieved August 11, 2009.
- ↑ ۵٫۰ ۵٫۱ Michael Ashburner, Golic KG, Hawley RS (2005). Drosophila: A Laboratory Handbook (2nd ed.). Cold Spring Harbor Laboratory Press. pp. 162–4. ISBN 0-87969-706-7.
{{cite book}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Bloomington Drosophila Stock Center at Indiana University (Bloomington): Basic Methods of Culturing Drosophila بایگانیشده در ۱ سپتامبر ۲۰۰۶ توسط Wayback Machine
- ↑ ۷٫۰ ۷٫۱ T.A. Brown (2007), GENOMES 3 (به انگلیسی), Garland Scienc Publishing
- ↑ Adams, M. D.; Celniker, S. E.; Holt, R. A.; Evans, C. A.; Gocayne, J. D.; Amanatides, P. G.; Scherer, S. E.; Li, P. W.; Hoskins, R. A. (2000-03-24). "The genome sequence of Drosophila melanogaster". Science (New York, N.Y.). 287 (5461): 2185–2195. doi:10.1126/science.287.5461.2185. ISSN 0036-8075. PMID 10731132.
- ↑ ۹٫۰ ۹٫۱ احمد مجد و محمدعلی شریعتزاده، زیستشناسی سلولی و ملکولی مجد، نشر آییژ
- ↑ Hartwell, Leland; Leroy Hood; Michael L. Goldberg; Ann E. Reynolds; Lee M. Silver (2011). Genetics:From Genes to Genomes; Fourth Edition. New York, NY: McGraw-Hill. ISBN 978-0-07-352526-6.
- ↑ Gilbert, Scott F. (2014). Developmental Biology, 10th edition. Sunderland, MA USA: Sinauer Associates, Inc. Publishers. p. 32.
- ↑ William D. Stansfield (۱۹۹۱), Schaum Outline of Theory and Problems of Genetics (به انگلیسی), McGraw-hill
- ↑ Reiter, LT; Potocki, L; Chien, S; Gribskov, M; Bier, E (2001). "A Systematic Analysis of Human Disease-Associated Gene Sequences In Drosophila melanogaster". Genome Research. 11 (6): 1114–1125. doi:10.1101/gr.169101. PMC 311089. PMID 11381037.
- ↑ Chien, Samson; Reiter, Lawrence T.; Bier, Ethan; Gribskov, Michael (1 January 2002). "Homophila: human disease gene cognates in Drosophila". Nucleic Acids Research. United States National Library of Medicine (NLM). 30 (1): 149–151. doi:10.1093/nar/30.1.149. PMC 99119. PMID 11752278. Retrieved August 24, 2013.
- ↑ Azpiazu N, Frasch M (1993). "tinman and bagpipe: two homeo box genes that determine cell fates in the dorsal mesoderm of Drosophila". Genes and Development. 7 (7b): 1325–1340. doi:10.1101/gad.7.7b.1325. PMID 8101173.
- ↑ فرهاد مشایخی (۱۳۹۰)، جنینشناسی، دانشگاه گیلان
- ↑ "Home Food Safety - Penn State Extension". extension.psu.edu (به انگلیسی). Retrieved 2024-07-20.